SAMPLE LAB REPORT
The Optimal Foraging Theory:
Food Selection in Beavers Based on Tree Species, Size, and Distance
Laboratory 1, Ecology 201
Abstract.
The theory of optimal foraging and its relation to central foraging
was examined by using the beaver as a model. Beaver food choice was
examined by noting the species of woody vegetation, status (chewed vs.
not-chewed), distance from the water, and circumference of trees near
a beaver pond in North Carolina. Beavers avoided certain species of
trees and preferred trees that were close to the water. No preference
for tree circumference was noted. These data suggest that beaver food
choice concurs with the optimal foraging theory.
Introduction
In this lab, we explore the theory
of optimal foraging and the theory of central place foraging using beavers
as the model animal. Foraging refers to the mammalian behavior associated
with searching for food. The optimal foraging theory assumes that animals
feed in a way that maximizes their net rate of energy intake per unit
time (Pyke et al. 1977). An animal may either maximize its daily energy
intake (energy maximizer) or minimize the time spent feeding (time minimizer)
in order to meet minimum requirements. Herbivores commonly behave as
energy maximizers (Belovsky 1986) and accomplish this maximizing behavior
by choosing food that is of high quality and has low-search and low-handling
time (Pyke et al. 1977).
The central
place theory is used to describe animals that collect food and store
it in a fixed location in their home range, the central place (Jenkins
1980). The factors associated with the optimal foraging theory also
apply to the central place theory. The central place theory predicts
that retrieval costs increase linearly with distance of the resource
from the central place (Rockwood and Hubbell 1987). Central place feeders
are very selective when choosing food that is far from the central place
since they have to spend time and energy hauling it back to the storage
site (Schoener 1979).
The main
objective of this lab was to determine beaver (Castor canadensis)
food selection based on tree species, size, and distance. Since beavers
are energy maximizers (Jenkins 1980, Belovsky 1984) and central place
feeders (McGinley and Whitam 1985), they make an excellent test animal
for the optimal foraging theory. Beavers eat several kinds of herbaceous
plants as well as the leaves, twigs, and bark of most species of woody
plants that grow near water (Jenkins and Busher 1979). By examining
the trees that are chewed or not-chewed in the beavers' home range,
an accurate assessment of food preferences among tree species may be
gained (Jenkins 1975). The purpose of this lab was to learn about the
optimal foraging theory. We wanted to know if beavers put the optimal
foraging theory into action when selecting food.
We hypothesized
that the beavers in this study will choose trees that are small in circumference
and closest to the water. Since the energy yield of tree species may
vary significantly, we also hypothesized that beavers will show a preference
for some species of trees over others regardless of circumference size
or distance from the central area. The optimal foraging theory and central
place theory lead us to predict that beavers, like most herbivores,
will maximize their net rate of energy intake per unit time. In order
to maximize energy, beavers will choose trees that are closest to their
central place (the water) and require the least retrieval cost. Since
beavers are trying to maximize energy, we hypothesized that they will
tend to select some species of trees over others on the basis of nutritional
value.
Methods
This study was conducted at Yates
Mill Pond, a research area owned by the North Carolina State University,
on October 25th, 1996. Our research area was located along
the edge of the pond and was approximately 100 m in length and 28 m
in width. There was no beaver activity observed beyond this width. The
circumference, the species, status (chewed or not- chewed), and distance
from the water were recorded for each tree in the study area. Due to
the large number of trees sampled, the work was evenly divided among
four groups of students working in quadrants. Each group contributed
to the overall data collected.
We conducted a chi-squared test
to analyze the data with respect to beaver selection of certain tree
species. We conducted t-tests to determine (1) if avoided trees were
significantly farther from the water than selected trees, and (2) if
chewed trees were significantly larger or smaller than not chewed trees.
Mean tree distance from the water and mean tree circumference were also
recorded.
Results
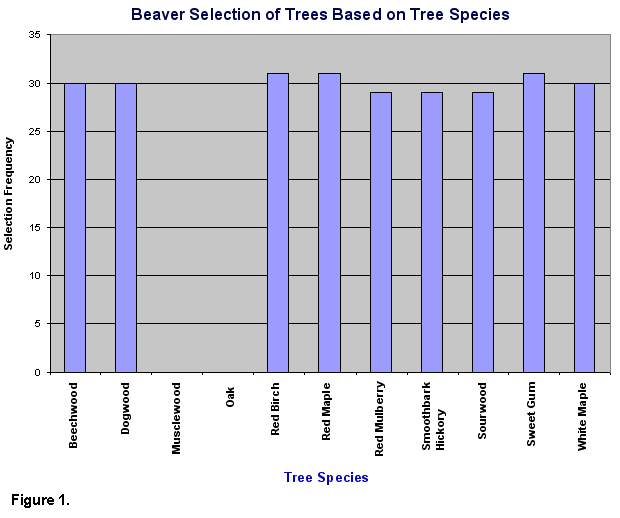
Overall, beavers showed a preference
for certain species of trees, and their preference was based on distance
from the central place. Measurements taken at the study site show that
beavers avoided oaks and musclewood (Fig. 1) and show a significant
food preference (x2=447.26, d.f.=9, P<.05). No avoidance
or particular preference was observed for the other tree species. The
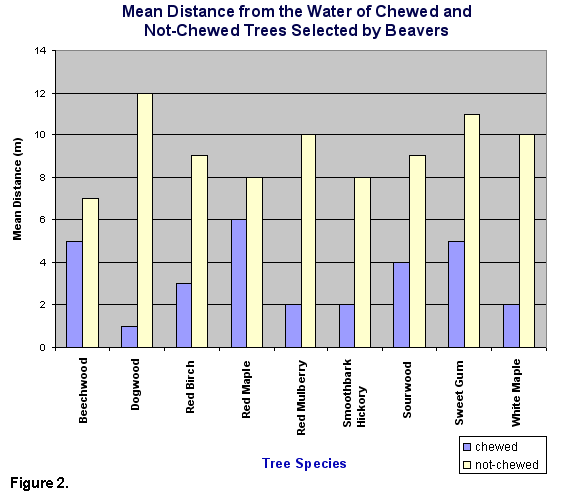
mean distance of 8.42 m away from the water for not-chewed trees was
significantly greater than the mean distance of 6.13 m for chewed trees
(t=3.49, d.f.=268, P<.05) (Fig. 2). The tree species that were avoided
were not significantly farther from the water (t=.4277, d.f.=268, P>.05)
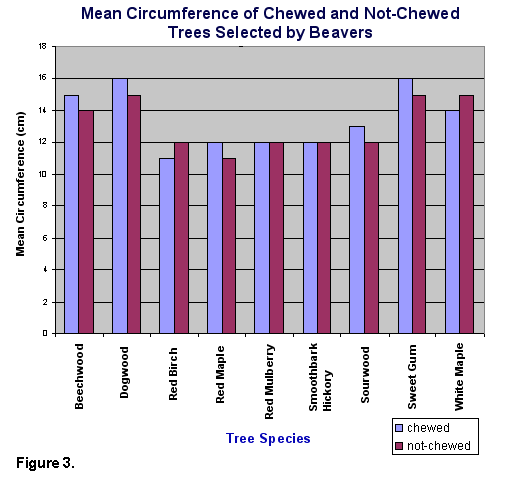
than selected trees. For the selected tree species, no significant difference
in circumference was found between trees that were not chewed (mean=16.03
cm) and chewed (mean=12.80 cm) (t=1.52, d.f.=268, P>.05) (Fig. 3).
Discussion
Although beavers are described
as generalized herbivores, the finding in this study related to species
selection suggests that beavers are selective in their food choice.
This finding agrees with our hypothesis that beavers are likely to show
a preference for certain tree species. Although beaver selection of
certain species of trees may be related to the nutritional value, additional
information is needed to determine why beavers select some tree species
over others. Other studies suggested that beavers avoid trees that have
chemical defenses that make the tree unpalatable to beavers (Muller-Schawarze
et al. 1994). These studies also suggested that beavers prefer trees
with soft wood, which could possibly explain the observed avoidance
of musclewood and oak in our study.
The result that chewed trees were
closer to the water accounts for the time and energy spent gathering
and hauling. This is in accordance with the optimal foraging theory
and agrees with our hypothesis that beavers will choose trees that are
close to the water. As distance from the water increases, a tree's net
energy yield decreases because food that is farther away is more likely
to increase search and retrieval time. This finding is similar to Belovskyís
finding of an inverse relationship between distance from the water and
percentage of plants cut.
The lack of any observed difference
in mean circumference between chewed and not chewed trees does not agree
with our hypothesis that beavers will prefer smaller trees to larger
ones. Our hypothesis was based on the idea that branches from smaller
trees will require less energy to cut and haul than those from larger
trees. Our finding is in accordance with other studies (Schoener 1979),
which have suggested that the value of all trees should decrease with
distance from the water but that beavers would benefit from choosing
large branches from large trees at all distances. This would explain
why there was no significant difference in circumference between chewed
and not-chewed trees.
This lab gave us the opportunity
to observe how a specific mammal selects foods that maximize energy
gains in accordance with the optimal foraging theory. Although beavers
adhere to the optimal foraging theory, without additional information
on relative nutritional value of tree species and the time and energy
costs of cutting certain tree species, no optimal diet predictions may
be made. Other information is also needed about predatory risk and its
role in food selection. Also, due to the large number of students taking
samples in the field, there may have been errors which may have affected
the accuracy and precision of our measurements. In order to corroborate
our findings, we suggest that this study be repeated by others.
Conclusion
The purpose of this lab was to
learn about the optimal foraging theory by measuring tree selection
in beavers. We now know that the optimal foraging theory allows us to
predict food-seeking behavior in beavers with respect to distance from
their central place and, to a certain extent, to variations in tree
species. We also learned that foraging behaviors and food selection
is not always straightforward. For instance, beavers selected large
branches at any distance from the water even though cutting large branches
may increase energy requirements. There seems to be a fine line between
energy intake and energy expenditure in beavers that is not so easily
predicted by any given theory.
Literature Cited
Belovsky, G.E. 1984. Summer diet
optimization by beaver. The American Midland Naturalist. 111: 209-222.
Belovsky, G.E. 1986. Optimal foraging
and community structure: implications for a guild of generalist grassland
herbivores. Oecologia. 70: 35-52.
Jenkins, S.H. 1975. Food selection
by beavers:› a multidimensional contingency table analysis. Oecologia.
21: 157-173.
Jenkins, S.H. 1980. A size-distance
relation in food selection by beavers. Ecology. 61: 740-746.
Jenkins, S.H., and P.E. Busher.
1979. Castor canadensis. Mammalian Species. 120: 1-8.
McGinly, M.A., and T.G. Whitham.
1985. Central place foraging by beavers (Castor Canadensis):
a test of foraging predictions and the impact of selective feeding on
the growth form of cottonwoods (Populus fremontii). Oecologia.
66: 558-562.
Muller-Schwarze, B.A. Schulte,
L. Sun, A. Muller-Schhwarze, and C. Muller-Schwarze. 1994. Red Maple
(Acer rubrum) inhibits feeding behavior by beaver (Castor
canadensis). Journal of Chemical Ecology. 20: 2021-2033.
Pyke, G.H., H.R. Pulliman, E.L.
Charnov. 1977. Optimal foraging. The Quarterly Review of Biology. 52:
137-154.
Rockwood, L.L., and S.P. Hubbell.
1987. Host-plant selection, diet diversity, and optimal foraging in
a tropical leaf-cutting ant. Oecologia. 74: 55-61.
Schoener, T.W. 1979. Generality
of the size-distance relation in models of optimal feeding. The American
Naturalist. 114: 902-912.
*Note: This document was modified from the work of Selena
Bauer, Miriam Ferzli, and Vanessa Sorensen, NCSU.
|